
Proper offshore farm design requires a framework capable of capturing the complex interplay between currents, surface waves, turbulence, canopy architecture, nutrient and light fields, and biological processes in an ever-changing ocean environment. To meet this challenge, MACMODS is a computational framework that integrates a state-of-the-art, open-source regional ocean model with a fine-scale hydrodynamic model. This model is capable of resolving turbulent fluxes within a canopy at sub-meter resolution and features a macroalgal growth model that accounts for hydrodynamically-mediated biological processes like the enhancement of nutrient uptake due to within canopy flows, plant motion, and waves. These tools are key to evaluate farm designs that maximize yield yet minimize cost and environmental impacts.
There are three basic components of MACMODS:
- Regional Ocean Model with biogeochemical fields (ROMS/BEC)
- Farm-scale hydrodynamic model (large eddy simulation, LES)
- Macroalgae Growth Model (MAG).
REGIONAL OCEAN CONDITIONS
ROMS/BEC
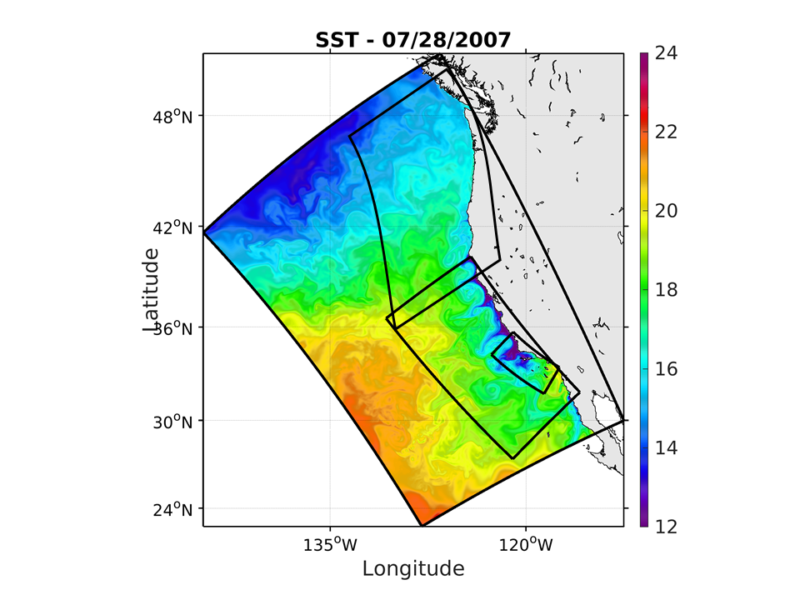
The Regional Modeling System (ROMS) is a widely-used, open-sourced code, which has been used for more than a decade in many locations in U.S. coastal waters and elsewhere (www.myroms.org; Shchepetkin and McWilliams 2005). Ecosystem and biogeochemical dynamics are predicted using the Biogeochemical Elemental Cycling (BEC) model (Moore et al., 2002; Gruber et al, 2006), run online embedded in ROMS. BEC includes phytoplankton, zooplankton, dissolved, suspended, and sinking particulates, including four different nutrients (nitrogen, silicic acid, phosphate, and iron). The model predicts carbonate system parameters, including pH, carbonate saturation, and pCO2, including air-sea exchanges. Recent improvements to the ROMS-BEC model of the Southern California Bight (SCB) include the addition of anthropogenic sources of nutrients from wastewater outfalls and riverine sources, which double the estimated amount of available nitrogen in the nearshore of the SCB (Howard et al. 2014).

FARM-SCALE HYDRODYNAMICS
WITHIN-FARM FLOWS
Cultivating offshore in deeper water requires macroalgae to grow from suspended structures, giving rise to a “suspended” canopy type (e.g., see Plew 2011). This is in contrast to many macroalgae types that are attached to the bottom, or extend from the bottom to the surface of the ocean (“submerged” and “emergent” canopy types). This new, emergent-type canopy will modify the interaction of hydrodynamics with the macroalgae farm. We implement a fine-scale turbulence model to assess how flow attributes, hydrodynamics forces, and nutrient and light availability respond to different farm designs and cultivation techniques.
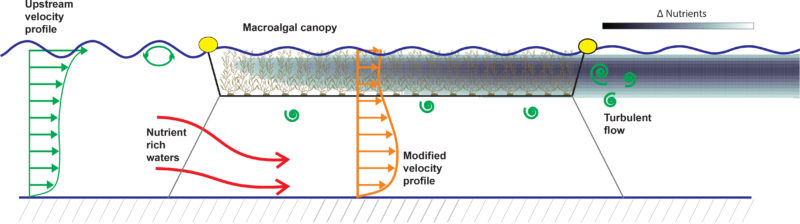
Large Eddy Simulation (LES) is the ideal approach for this investigation because it permits accurate representation of the turbulent fluid motions responsible for most of the drag forces exerted on plants and farm structures, and for the mass/nutrient transport within the canopy. In LES, one can capture the modulation of turbulence by the mean wave effect and the formation of Langmuir cells and the development of shear layer eddies. The LES model consists of three main components: (i) a dynamical core to calculate the fluid motion including those induced by winds, waves, density stratification, and bottom interactions, (ii) a canopy module to represent the effects of the macroalgal canopy and plant motion on the flow field, and (iii) a scalar module to represent the transport of nutrients through the canopy.
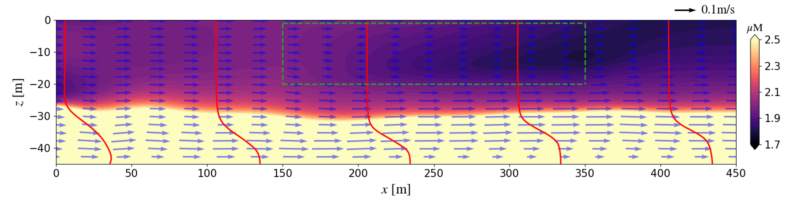
Movie shows velocity vectors and nutrient concentrations (color contours) for an LES run of a Macrocystis pyrifera (giant kelp) farm (farm canopy is in the green box) located on the San Pedro shelf (ROMS boundary conditions). Note how the flow slows in the farm due to canopy drag, but that nutrients are entrained upward into the canopy by turbulent mixing.
MACROALGAE GROWTH MODEL
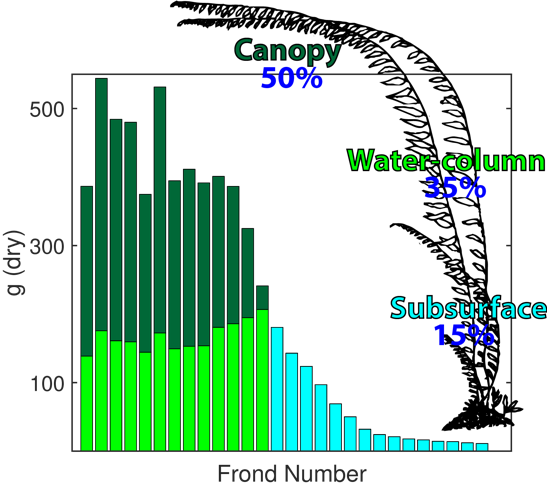
The MACROALGAE GROWTH MODEL (MAG) has been designed to be flexible and easily adapted to many types of macroalgae. Initially we have developed and parameterized MAG for giant kelp, Macrocystis pyrifera. The model tracks nitrogen in six pools: ammonium, nitrate, dissolved organic nitrogen, particulate organic nitrogen, stored nitrogen in macroalgae, and fixed nitrogen in macroalgae. The currency of nitrogen has been selected as this is most often the nutrient limiting autotroph productivity along the U.S. West Coast (Eppley et al. 1979), including for kelp (Zimmerman and Kremer 1984, 1986).

References:
Gruber, N., Frenzel, H., Doney, S. C., Marchesiello, P. Mcwilliams, J. C., Mosian, J. R., Oram, J. J., Plattner, G. & Stolzenbach K. D. (2006). Eddy-resolving simulation of plankton ecosystem dynamics in the California Current System. Deep Sea Research Part I Oceanographic Research Paper, 53 (9): 1483-1516.
Howard, M. D., Sutula, M., Caron, D. A., Chao, Y., Farrara, J. D., Frenzel, H., Burton, J., George, R., MaLaughlin, K. & Sengupta, A. (2014). Anthropogenic nutrient sources rival natural sources on small scales in the coastal waters of the Southern California Bight. Limnology and Oceanography.
Moore, J. K., Doney, S. C., Kleypas, J. A., Glover, D. M., & Fung, I. Y. (2001). An intermediate complexity marine ecosystem model for the global domain. Deep Sea Research Part II Topical Studies in Oceanography, 49(1), 403-462.
Shchepetkin, A. F., & McWilliams, J. C. (2005). The regional oceanic modeling system (ROMS): a split-explicit, free-surface, topography-following-coordinate oceanic model. Ocean Modelling, 9(4), 347-404.
Eppley, R. W., Renger, E. H., Harrison, W. G., & Cullen, J. J. (1979). Ammonium distribution in southern California coastal waters and its role in the growth of phytoplankton. Limnology and Oceanography, 24(3), 495-509.
Zimmerman, R. C. & Kremer, J. N. (1984). Episodic nutrient supply to a kelp forest ecosystem in Southern California. Journal of Marine Research, 42: 591–604.
Zimmerman, R. C., & Kremer, J. N. (1986). In situ growth and chemical composition of the giant kelp, Macrocystis pyrifera: response to temporal changes in ambient nutrient availability. Marine Ecology Progress Series, 27(3).